Immune Checkpoint Blockers Deliver Success in Cancer Immunotherapy
Summary
Monoclonal antibodies such as ipilimumab and pembrolizumab that block immune checkpoints have delivered spectacular results in the clinic and promise a revolution in cancer therapy. Here we review their history, fundamental mechanism of action, and highlight some of the pipeline products and future prospects of this “game-changing” class of drugs.
Historical Background
The recent emergence of cancer immunotherapy, named as “Breakthrough of the Year” in 2013 by Science magazine, belies its origins which lie with William Coley, an American bone surgeon born in 1867. Coley (pictured left) made the connection between the immune system and cancer, and developed a theory that post-surgical infections helped patients recover from bone cancer by provoking an immune response. He even treated cancer patients with “Coley’s toxins”, a crude bacterial extract, claiming successful treatment of hundreds of patients. However, with unpredictable results, low success rates and the emergence of other treatments such as radiotherapy, an immune based approach was neglected. It would take decades of basic research unravelling the complexities of the immune system, in particular the contribution of different cell types, to inform and enable more sophisticated interventions, and return the focus to cancer immunotherapy.
T Cell Activation Requires (at least) Two Signals
We now know that white blood cells called T lymphocytes (T cells) are central to immune responses, and without T cells (for example in untreated HIV infected individuals) the immune system simply doesn’t work. T cells become activated by antigen presenting cells (APC’s) such as dendritic cells, and determine the specificity of immune responses since they only react to particular antigens that are recognised by the T cell antigen receptor (TcR).
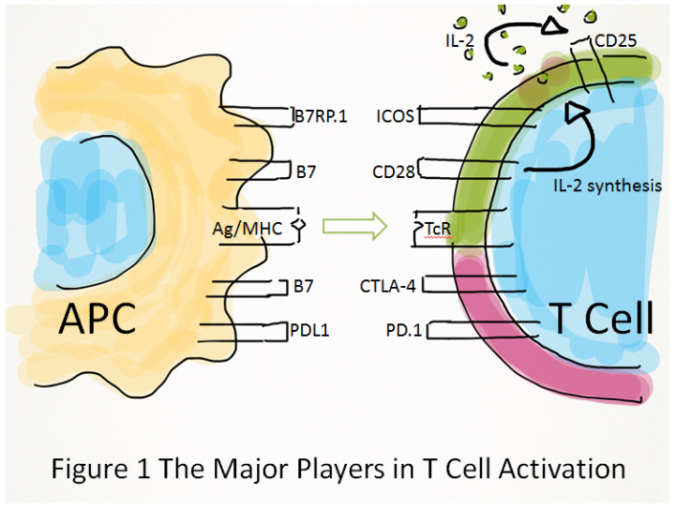
T cell activation is initiated by antigens in combination with major histocompatibility proteins (Ag/MHC), on the surface of antigen presenting cells (APC’s). These bind (green arrow) the T cell antigen receptor (TcR) to propagate intracellular signals in the T cell). Full T cell activation requires a second “co-stimulatory” signal (indicated by green T cell plasma membrane) for example CD28 (which drives synthesis of the T cell growth factor interleukin-2, IL-2) or ICOS signals. CD25 is the IL-2 receptor. Some receptors however serve to limit T cell activation (indicated by red plasma membrane), e.g. CTLA-4 and PD-1. These are known as “immune checkpoints” which can be blocked by monoclonal antibodies to boost otherwise weak anti-tumour immune responses.
Intracellular signals propagated by the TcR alone however, are not sufficient to fully activate a T cell but a second signal called “co-stimulation” is required (represented in Figure 1 above). The first receptor identified that could co-stimulate T cells was cluster of differentiation 28 (CD28). B7 is the ligand (binding partner) for CD28, and is expressed on the surface of APC’s. When bound by B7, CD28 stimulates the production of interleukin-2 (IL-2), a growth factor that drives T cell proliferation, “arming” the body with a sufficient mass of antigen specific T cells required for a meaningful immune response. The profound effect of co-stimulation on the subsequent course of immune responses makes the T cell/ APC interface a key target for immunomodulatory drugs.
Development Stage of Some Immune Checkpoint Blocking Drugs
| Target | Biological Function | Drug | Development Stage |
|---|---|---|---|
| CTLA-4 | Inhibitory receptor | Ipilimumab | Approved for melanoma, Phase II and III trials for multiple cancers |
| Tremelimumab | Previously tested in Phase II trial for melanoma | ||
| PD1 | Inhibitory receptor | Nivolumab(MDX-1106) | Phase I/II trials in patients with melanoma, renal and lung cancers |
| Pembrolizumab(MK3475) | Phase I trials in multiple cancers | ||
| Pidilizumab(CT-011) | Phase II trials in diffuse large B cell lymphoma, Phase I trials in multiple cancers | ||
| AMP224 | Phase I trials in cancer | ||
| PDL1 | Ligand for PD1 | MDX-1105 | Phase I trials in multiple cancers |
| LAG3 | Inhibitory receptor | IMP321 | Phase III trials in breast cancer |
| B7-H3 | Inhibitory ligand | MGA-271 | Phase I trials in multiple cancers |
| B7-H4 TIM3 | Inhibitory ligand | Pre-clinical development | |
| CTLA-4, cytotoxic T-lymphocyte antigen-4. CD28, cluster of differentiation 28. PD1, programmed death protein 1. LAG3, lymphocyte activation gene 3, TIM3, T cell membrane protein-3. B7-H3 and B7-H4 ligands have not yet been identified, but are presumed to function as T cell inhibitory receptors from gene knock out experiments. | |||
CTLA-4: CD28’s “Evil Twin” A second B7 ligand expressed on T cells was identified in cytotoxic T-lymphocyte antigen-4 (CTLA-4, also known as CD152) that had structural and signalling similarities to CD28. Functionally however CTLA-4 is opposed to CD28, preventing T cell activation and limiting immune responses, hence it is known as an “immune checkpoint” (see Figure 1). It is thought that without checkpoints like CTLA-4, immune responses could cause damage to self-tissues resulting in auto-immune inflammatory diseases such as arthritis. However, blocking immune checkpoints by using monoclonal antibodies to prevent receptor-ligand interactions can be beneficial to cancer patients by boosting otherwise weak anti-tumour responses. This is the basic mechanism, of action of the immune checkpoint blockers.
An array of positive and negative regulators of T cell activation have now been identified, summarised as “target” in the table above, indeed the receptors targeted clinically so far represent only a fraction of potential candidates. The clinical development stage of therapeutic proteins targeting immune checkpoints is outlined in the table is illustrative rather than exhaustive. Since receptors that function as immune checkpoints differ in their signalling and expression patterns, it can be assumed that they are also functionally different. For example, in contrast to CTLA-4 it appears the role of PD-1 is to limit the activity of T cells in peripheral tissues. This raises the possibility that blocking multiple checkpoints may yield additive or even synergistic results.
The Specific/Non-specific Paradox
Monoclonal antibody based therapies exploit the exquisite specificity of protein-protein interactions to minimise “off-target” effects. This benefits the patient by limiting the side effects of drugs that can be gruelling, for example in chemotherapeutic regimes. However, the receptors that are targeted by immune checkpoint blockers show no inherent tumour specificity. Initially this lead some to question the strategy of targeting CTLA-4 however, now the approach has been validated in the clinic it presents an opportunity to apply the approach to cancers beyond those already tested.
Future Prospects/Questions
The success of immune checkpoint blockers in the clinic immediately gives rise to a number of questions such as follows:
- Are there other immune check points that can be exploited therapeutically?
- Which other cancers can be treated in this way?
- Are there biomarkers that indicate which check point is most relevant to a particular cancer?
- Can combination therapies targeting multiple checkpoints yield yet more effective results?
- Are some checkpoint blockers/combinations more effective in particular cancers?
- Can the use of combinations of checkpoint blockers minimise drug side effects?
Answering these questions will involve years of work in clinical trials, but holds the promise of delivering yet more “weapons” to physicians in the fight against cancer. The translation of basic research of T cell physiology and function into new therapeutic strategies has taken decades of hard work but is now a major success story. The potential pipeline of drugs, coupled with the refinement in the way they are used, could return the focus to immune based therapies in the treatment of cancer, ultimately to the benefit of patients.